草魚呼腸孤病毒培養與滴度測定、及在草魚、CIK細胞上的生長特性研究(三)
2.2 GCRV 873株在CIK細胞中的生長特性
GCRV 873株吸附CIK細胞1 h后,分別在0、2、4、8、10、12、24、36、48、72 h收取細胞病毒液。對各個時間點病毒滴度及RNA水平進行檢測(圖3),并通過顯微鏡觀察不同時間點細胞病變情況(圖4)。由圖3可以看出病毒在感染CIK細胞0~12 h期間,病毒RNA水平很低,上升不明顯,病毒滴度處于較低水平,上升趨勢也不明顯;12~24 h期間,病毒RNA和病毒滴度開始明顯上升;24~72 h期間,病毒RNA急劇上升,而病毒滴度在24~48 h期間明顯上升,48 h后趨于平穩,72 h時病毒滴度達到最高,為106.75TCID50/mL。GCRV 873株感染CIK后12 h時出現少量細胞聚集;24 h時,細胞聚集增多,出現空斑現象;36 h時,空斑變大;48 h時,細胞大量脫落;72 h時,細胞基本已脫落,只有零星細胞附著于板上(圖5)。
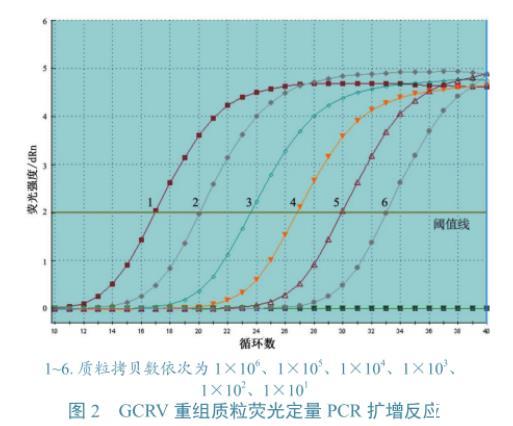
圖2 GCRV重組質粒熒光定量PCR擴增反應
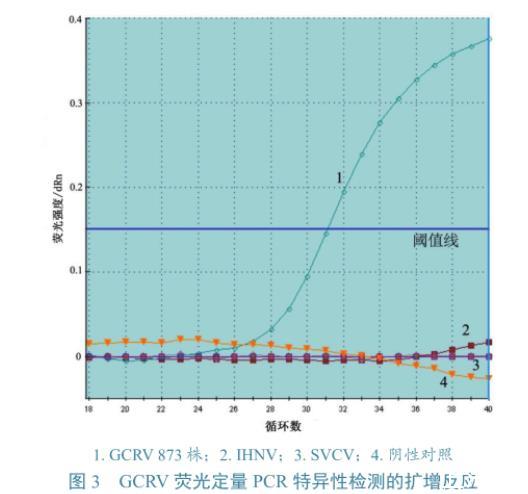
圖3 GCRV熒光定量PCR特異性檢測的擴增反應
2.3 GCRV 873株在草魚體內的復制情況
GCRV 873株感染草魚后,其肝、脾、腎中的病毒RNA拷貝數變化如圖6所示。3個組織中的病毒RNA均呈先升后降趨勢,其中肝臟中的病毒RNA峰值位于感染后3 d,脾臟和腎臟峰值位于感染后2 d。3種組織中,病毒RNA峰值最高的為腎臟,肝臟次之,脾臟最低;肝臟、腎臟和脾臟中的病毒RNA分別在至感染后7、6、4 d時下降到趨近于零的水平。
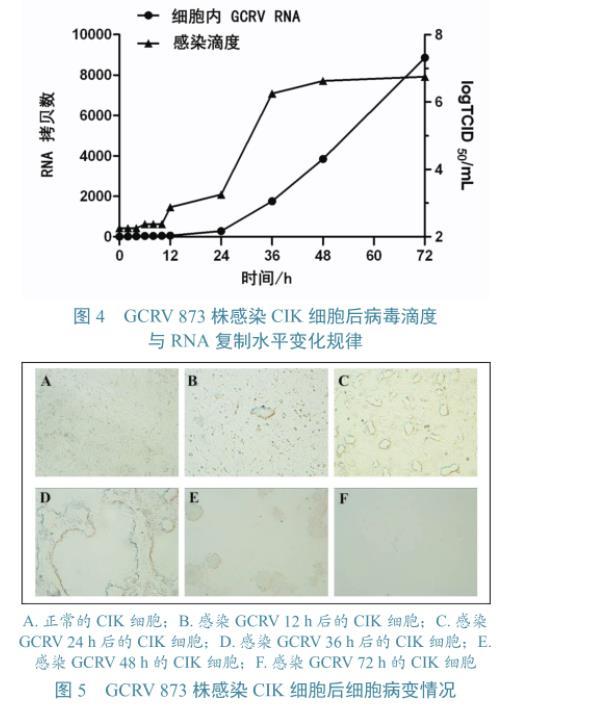
圖4 GCRV 873株感染CIK細胞后病毒滴度與RNA復制水平變化規律 圖5 GCRV 873株感染CIK細胞后細胞病變情況
3討論
本研究從病毒RNA拷貝數、子代病毒含量及細胞病變3個方面綜合分析GCRV在CIK細胞中的增殖過程。試驗結果表明,GCRV在感染CIK細胞的早期病毒RNA拷貝數及病毒滴度均處于較低水平,說明這一時期的病毒多為殘余的原代病毒及最先釋放出來的子代病毒;24~72 h期間,病毒RNA水平迅速上調,呈現對數增長,而病毒滴度在24~48 h期間呈明顯上升,之后趨于平穩;對照48 h的細胞CPE發展完全,說明細胞被完全裂解,胞內病毒得以釋放,使病毒滴度趨于平穩。曾令兵等利用組織培養微量滴定系統,研究GCRV-854毒株在CIK細胞上的繁殖過程,其動態曲線與本研究相似,但出現快速增殖的時間較本研究晚,說明不同毒株感染細胞的能力存在差異。鄒桂平等在電鏡下觀察到GCRV在感染CIK細胞4 h以內出現脫去部分外層衣殼的不完整病毒顆粒,感染8 h時漿胞內出現大量亞病毒顆粒,無外層蛋白結構,感染12~16 h后出現成熟的病毒粒子。丁清泉等發現GCRV在感染細胞12 h后即開始增殖,24~72 h大量增殖,使細胞產生典型的細胞病變效應,5 d左右達到最大增殖,此時病毒的滴度最高,以后逐漸平緩。分析病毒在細胞中的增殖過程,不僅反映出病毒的生長特性,還指示出收獲細胞培養的病毒材料的最佳時間。
應用Real-time PCR技術,實時監測GCRV在草魚體內的病毒拷貝數,為精確分析病毒在魚體中的增殖過程提供基礎數據。本試驗采用的毒株GCRV 873株為1型。該毒株在體外細胞中培養可使細胞產生明顯病變,但是草魚感染后未出現發病死亡情況,而其肝、脾、腎中的病毒RNA水平出現短期上升后下降到對照水平的結果,也正與無發病死亡情況相符。殷亮研究發現GCRV HZ08株在感染稀有鮈鯽后,脾臟和腎臟中的病毒RNA水平呈現先升后降趨勢,與本研究結果一致。丁清泉等發現經人工感染GCRV的魚體腎臟組織細胞內,存在無外衣殼的未成熟病毒。毛樹堅等觀察草魚出血病的病理切片發現在肝臟、肌肉、腎臟、脾臟、鰓等魚體組織中含有大量的病毒顆粒。在本研究對比3種組織中病毒RNA的含量,發現腎臟中的含量最高,提示在取樣檢測時,可優先考慮腎臟組織。
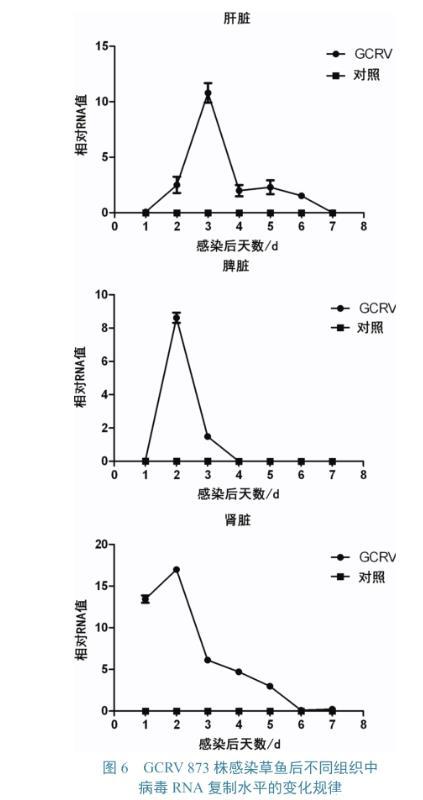
圖6 GCRV 873株感染草魚后不同組織中病毒RNA復制水平的變化規律
4結論
本研究顯示:GCRV在感染CIK細胞12 h后病毒RNA和病毒滴度開始上升;24~72 h期間,病毒RNA急劇上升,病毒滴度則在24~48 h內明顯上升,48 h后趨于平穩,至72 h,病毒滴度達到最高,為106.75TCID50/mL。在感染CIK后36 h,所有細胞均已感染,72 h時,細胞基本上脫落。GCRV感染草魚后,均能在其肝、脾、腎中檢測到病毒,且病毒RNA均呈先升后降趨勢;腎臟中的病毒RNA含量最高,因此應優先采集腎臟組織進行檢測。
相關新聞推薦
1、不同碳源、氮源和金屬離子對耐鋅菌株的生長特性的影響(二)
3、稻花香馫香型根霉曲、中高溫大曲和高溫大曲的酶活力和微生物數量統計(上)